Центромеры
Оглавление.
Центромеры дрожжей
Центромеры человека
Центромеры дрозофилы
Центромеры арабидопсиса
Reference
Центромеры - это хромосомные структуры ответственные за направление движения хромосом во время митоза. К функциям центромер относятся адгезия сестринских хроматид, образование кинетохора, спаривание гомологичных хромосом и вовлечение в контроль генетической экспрессии. У большинства эукариот центромеры не содержат определенной последовательности ДНК. Обычно они содержат повторы (например, сателлитной ДНК), схожие, но не идентичные. У нематоды Caenorhabditis elegans и некоторых растений хромосомы голоцентрические, т.е. образование кинетохора не локализовано определенным участком, а происходит диффузно по всей длине хромосомы.
Центромеры дрожжей.Центромера Sp длинной 35-110 тпн (чем хромосома длиннее, тем центромера меньше) и состоит из двух доменов - центральной коровой области и внешней области повторов (otr), предстравленной гетерохроматином (рис1). Центральная коровая область состоит из области неповторяющейся ДНК (cnt) и области инвертированных повторов (imt) по краям cnt. В центральной коровой области нормальный гистон H3 заменен своим аналогом (CENP-A у Sc) и в этом месте собирается кинетохор. Маркерные гены встраеваемые в центромерную последовательность становятся транскрипционно неактивными. Их замолкание зависит от положения, например, на внешних повторах оно сильнее, а в центральной области менее выражено. Белки Mis6, Mis12, Mal2 и Sim4 связываются с центральным районом центромеры. Центральный район частично переваривается микрококковой нуклеазой, что указывает на особую организацию хроматина, причем эта организация не зависит от ДНК (ДНК перенесенная в Sp или в другие участки хромосомы не сохраняет такую организацию). Внешние повторы упакованы в нуклеосомы, с деацетилированными гистонами (при помощи деацетилаз Clr3, Clr6 и Sir2). Метилтрансфераза Clr4 диметилирует H3K9, на который садится Swi6 (аналог HP1) и Chp1. Таким образом, на центромере формируется гетерохроматин (см. обзор Гетерохроматин). Swi6 отвечает за присоединение когезинов к области внешних повторов. otr состоят из dg и dh повторов, разделенные другими повторами. Внутренние и внешние повторы содержат кластеры генов тРНК. Установлено, что dg повторы имеют первостепенную роль в установлении центромерной активности.
ДНК центральной коровой области АТ-богатая и состоит из трех участков cnt1, cnt3 - гомологичны на 99%, расположены по кроям от сnt3 гомологичного с ними на 48%. Левый и правый imr инвертированы и уникальны для каждой центромеры.
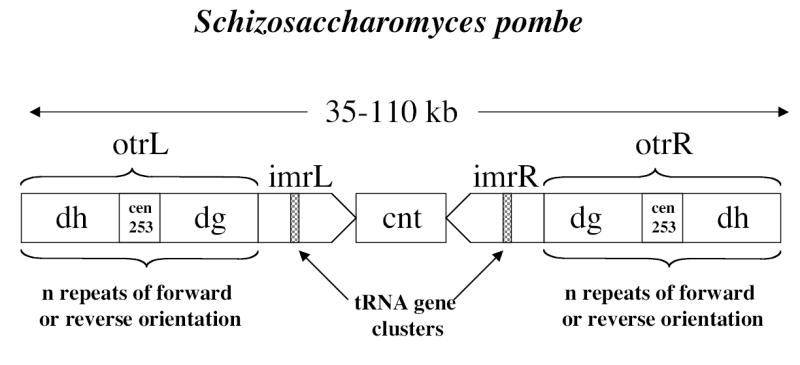
рис.1
Все 16 центромер Sc имеют длину 90 пн и содержат три элемента: CDEI, CDEII и CDEIII (рис.2). CDEII - это АТ-богатый неконсервативный спейсер длинной 78-90 пн, разделяющий CDEI и CDEIII. CDEI имеет длинну8 пн. Этот участок не существенен для центромерной активности, но его делеция повышает вероятность неправильного расхождения хромосом во время митоза. СDEII - 78-90 пн, содержит ~90% АТ-пар. Делеции в этом участке прерывают образование центромеры, не нарушая расхождение хромомсом. СDEIII - 26 пн содержит несовершенные палиндромы. Одиночная нуклеотидная замена в этом участке полностью прерывает центромерную активность.

рис.2
Центромеры человека
Центромера человека представляет участок 1-4 Мпн а-сателлита длинной 171
пн (альфоид). Другие сателлиты также присутствуют. В пределах повторов
устанавливается место образования центромеры называемое неоцентромера.
Первичная последовательность ДНК в установившейся неоцентромере не имеет
значения. Не все а-сателлиты становятся центромерой, не смотря на присутствие
двух локусой богатых а-сателлитом, активной центромерой становится только
один из них. Интактная ДНК, содержащая альфоид и помещенная в ядро, не
образует активной центромеры, поэтому первичный механизм образования активной
центромеры остается неясным.
Сокращения:
Sp (Schizosaccharomyces pombe ) - делящиеся дрожжи
Sc (Saccharomyces cerevisiae ) - почкующиеся дрожжи
cnt (central core region) - центральная коровая область
imr (innermost repeat) - внутренние повторы
otr (outer repeats) - внешние повторы
Suvar
June 5, 2006 22:09
Свои пожелания отправляйте по адресу
drosophila.narod.ru
Жизнь коротка, наука вечна
Наука есть лучший способ удовлетворения любопытства за счет государства.
(Л.А.Арцимович)
![]()