Контрольные точки клеточного цикла
Оглавление.
Участники контроля клеточного цикла
Контрольные точки клеточного цикла
Контроль различных этапов клеточного цикла
Циклин-зависимые киназы
Циклины
Субстраты циклинов
APC комплекс
Ингибиторы комплексов Cdk-циклин
Участники контроля клеточного цикла.
Клеточный цикл контролируется путем взаимодействия трех типов белков:
циклинзависимые киназы (Cdk), циклины - белки, взаимодействующие
с Cdk c образованием комплексов и ингибиторы комплексов Cdk-циклин.
Циклинзависимые киназы (Cdk) - ферменты фосфорилирующих другие белки,
изменяя их функцию. Клеточный цикл контролируется изменением активности
Cdk, которая регулируется периодическим образованием и распадом их регуляторных
субъединиц - циклинов. Смена синтезов и разрушений различных циклинов
обеспечивает переходы и протекания различных фаз клеточного цикла. При
этом концентрация Cdk постоянна в течении всего клеточного цикла. В разные
фазы клеточного цикла образуются разные циклины, которые связываясь с
Cdk образуют различные Cdk-циклиновые комплексы. Эти комплексы регулируют
разные фазы клеточного цикла и поэтому называются G1-, G1/S-
, S- и М-Cdk (рис.1).

рис.1 Концентрации различных комплексов Cdk-циклин в клеточном
цикле.
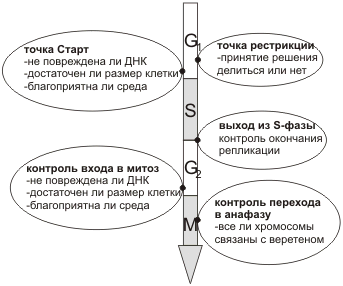
Контрольные точки клеточного цикла.
1. Точка выхода из G1-фазы, называемая Старт
- у млекопитающих и точкой рестрикции у дрожжей. После перехода через
точку рестрикции R в конце G1 наступление S становится необратимым,
т.е. запускаются процессы ведущие к следующему делению клетки.
2. Точка S – проверка точности репликации.
3. Точка G2/M-перехода – проверка завершения
репликации.
4. Переход от метафазы к анафазе митоза.
Контроль различных этапов клеточного цикла
ARC подавляет S- и M-циклины и не подавляет G1/S-циклины.
В G1-фазе работают различные ингибиторы Cdk.
Внутренние и внешнии сигналы приводят к образованию G1/S- и
S-циклинов и активации G1/S–Cdks.
Активность G1/S–Cdk увеличивается потому что G1/S циклины не атакуются
APC и потому что G1
Cdk ингибиторы так же не действуют на G1/S–Cdks (у мух и дрожжей)
или удаляются от G1/S–Cdks другими механизмами (у млекопитающих).
S-Cdk инактивирует ингибиторы Cdk и подавляет ARC, которые в G1-фазе
подавляли S-Cdk. S-Cdk фосфорилируют различные белки, что ведет к началу
дупликации ДНК и S-фазы. После начала S-фазы S/G1-Cdk обеспечивают
собственную инактивацию.
В конце S-фазы, в G2-фазе начинают накапливаться М-Cdk, приводящая
к вступлению клетки в митоз. М-Cdk активирует ARC-комплекс, управляющий
метафазно-анафазным переходом. Основная функция ARC-комплекса состоит
в разрушении когезинов, приводящее к началу расхождения хромосом
Циклин зависимые киназы Cdk1-5 в клетках млекопитающих
Cdks активируется при связывании с циклинами (так же как фосфориляция
и дефосфориляция киназ). Cdks-фосфорилируют белки участвующие в кл цикле
M-phase Cdk (M-Cdk) запускают каскад белковых фосфориляций, запускающих
М-фазу к.ц. (конденсация хромосом, разрушение ядра, перестройка АГ иЭР,
потеря адгезии с большинством других клеток и внеклеточному матриксу,
реорганизация цитоскелета)
anaphase-promoting complex (APC) регулятор митоза – инициация разделения
и расхождения хромосом и инактивация М-Cdk в конце митоза
При выходе из G0 под действием факторов роста начинает синтезироваться
Cdk2-циклинD: распознает в-ва, регулирующие ферменты синтеза белков, необходимых
для репликации ДНК. В это же время выявляются Cdk4-циклинD, и Cdk5циклинD
циклин-cdks запускает М-стадию кц, деградация циклина снижает активность
cdks
Cdk2-циклинE появляется в G1 и достигает max на границе G1-S, после чего
его концентрация резко снижается
Cdk2-циклинА появляется в промежутке G1-S и присутствует в высокой концентрации
на протяжении S
Сdk2-циклинB в конце G2 до М – резко разрушается
в каждой стадии синтезируются свои циклины M-циклины запускают события
митоза, G1/S-циклины – связывают цзк в конце G1
подготавливает кл к S-фазе, S-циклины – связывают цзк, запуская репликацию,
G1-циклины обеспечивают прохождение через точку рестрикции.
Регуляция репликации
Перед началом репликации Sc ORC-комплекс (origin recognition
complex) садится на ori - точку начала репликации. Cdc6 представлен во
всем клеточном цикле, но его концентрация возрастает вначале G1, где он
связывается c ОRC комплексом, к которому затем присоединяются Mcm белки
с образованием pre-replicative complex (pre-RC). После сборки pre-RC клетка
готова к репликации.
Для инициации репликации S-Cdk соединяется с протеинкиназой (?), которая
фосфорилирует pre-RC. При этом Cdc6 диссоциирует от ОRC после начала репликации
и фосфорилируется, после чего убиквитинируется SCF и деградирует. Изменения
в pre-RC препятствуют повторному запуску репликации. S-Cdk так же фосфорилирует
некоторые Mcm белковые комплексы, что запускает их экспорт из ядра. Последующая
дефосфориляция белков вновь запустит процесс образования pre-RC.
Регуляция митоза
В эмбриональных клетках синтез М-циклина постоянен во всем клеточном цикле
и накопление его происходит из-за уменьшения деградации. У большинства
клеток М-циклин синтезируется во время G2 и М-фаз. Накопление циклина
ведет к накоплению M-Cdk. Cdk ингибируется, фосфорилируясь протеинкиназой
Wee1. Активация Cdc25 в поздней G2 дефосфорилирует M-Cdk, так же происходит
репрессия Wee1. Две протеинкиназы фосфорилируют Cdc25 – Polo kinase и
M-Cdk. M-Cdk так же фосфорилирует и ингибирует Wee1. Способность M-Cdk
активировать свой собственный активатор (Cdc25) и ингибировать свой собственный
ингибитор (Wee1) предполагает, что активация M-Cdk в митозе резко усиливается
при наличии такой позитивной обратной связи. Малое количество активированных
Cdc25 активируют M-Cdk, которые активирует еще больше Cdc25 и супрессируют
Wee1. Это приводит к большей дефосфориляции M-Cdk и активации и тд. Такой
механизм обеспечивает полную активацию всех M-Cdk
Фосфорилирование ламинов M-Cdk приводит к их деградации. М-Cdk фосфорилирует
несколько субъединиц конденсинов, запуская конденсацию хромосом.
M-Cdk фосфорилирует различные белки, запуская реорганизацию микротрубочек
и другие события ведущие к организации веретена деления.
| вид | название | синоним | размер | функция |
| S.cerevisiae | Cdk1 | Cdc28 | 298 | все стадии клеточного цикла |
| S.pombe | Cdk1 | Cdc2 | 297 | все стадии клеточного цикла |
| D.melanogaster | Cdk1 |
Cdc2 Cdc2c Cdk4/6 |
297 314 317 |
M G1/S, S, возможно М G1 обеспечивает рост |
| X.laevis | Cdk1 Cdk2 |
Cdc2 |
301 297 |
M S, возможно S |
| H.sapiens | Cdk1 Cdk2 Cdk4 Cdk6 |
Cdc2 |
297 298 303 326 |
M G1/S, S, возможно М G1 G1 |
В животных клетках имеются, по крайней мере, 7 различных Cdk. Cdk1,2,4,6
напрямую участвуют в регуляции клеточного цикла, тогда как остальные фосфорилируют
другие Cdk и называются Cdk-активирующие киназы (CAK).
Cdk7,8,9 являются регуляторами РНК полимеразы II. Cdk5 участвует в дифференцировке
нервных клеток.
У дрожжей Sc и Sp все события клеточного цикла контролируются одной Cdk1.
У многоклеточных организмов события контролируются Cdk1 и Cdk2. Также
у высших эукариот имеются Cdk4 и Cdk6 которые регулируют клеточный цикл
в ответ на внеклеточные сигналы.
Cdk фосфорилируют сотни различных белков по сериновым (S) или треониновым
(T) аминокислотным остаткам. Cdk узнает мотиф другого белка по которому
необходимо фосфорилировать: [S/T*]PX[K/R], где S/T*- место фосфорилирования,
X – любая аминокислота, K/R-основные аминокислоты лизин (K) или аргинин
(R).
В отсутствии циклина активный центр Cdk заблокирован.
Cdk состоит из нескольких доменов: Т-петля (инактивирующая петля) – закрывает
активный центр в отсутствии циклина. L12 helix, PSTAIRE helix.
| Вид | G1 | G1/S | S | M |
| S.cerevisiae | Cln3 (Cdk1) |
Cln1,2 (Cdk1) |
Clb5,6 (Cdk1) |
Clb1,2,3,4 (Cdk1) |
| S.pombe | Puc1? (Cdk1) |
Puc1 Cig1?, (Cdk1) |
Cig2, Cig1? (Cdk1) |
Cdc13 (Cdk1) |
| D.melanogaster | cyclin D (Cdk4) |
cyclin E (Cdk2) |
cyclin E, A (Cdk2,1) |
cyclin A, B, B3 (Cdk1) |
| X.laevis | cyclin E (Cdk2) |
cyclin E, A (Cdk2,1) |
cyclin A, B (Cdk1) |
|
| H.sapiens | cyclin D1,2,3 (Cdk4,6) |
cyclin E (Cdk2) |
cyclin A (Cdk2,1) |
cyclin B (Cdk1) |
Циклины - цитоплазматические белки. Разрушение циклинов происходит в
протеосомах (см. обзор Протеасомы). Циклин B – белок киназный домен, регуляторная
субъединица. Начинает синтезироваться в G1, достигает max в S и ранней
профазе и быстро разрушается в начале анафазы М. Когда концентрация регуляторной
субъединицы возрастает – активируется киназный домен. Фосфорилирование
специфических белков приводит к компактизации х-м, разрушению ядерной
об-ки и сборке веретена.
Циклин разрушается в протеасомах. Сигнальная послед-ть из 8-10 а-кт –блок
разрушения - узнается распознающим белком, убиквитинлигаза присоединиет
убиквитин (76а-кт) к остаткам лизина, который полиUb и узнается РНК-белковым
комплексом протеасомой. Фосфатаза переводит распознающий белок в неактивное
состояние, а циклин фосфорилирует переводя в активное ?чем больше циклина,
тем быстрее он разрушается.
Циклин фосфорилирует сериновые и треониновые остатки ламинов вызывая их
деполимеризацию, фосфорилирует гистон H1, участвует в фосфорилировании
блокирующим везикулярный транспорт – разрушение ЭПР и АГ, фосфорилирует
участок легкой цепи миозина, ингибируя АТФ-азную активность и связывание
с F-актином – блокировка цитокинеза в раннем митозе. После разрушения
циклина белки дефосфорилируются.
Циклины – активаторы Cdk. Циклины, так же как и Cdk вовлечены в различные,
помимо контроля клеточного цикла, процессы. Циклины разделяются на 4 класса
в зависимости от времени действия в клеточном цикле: G1/S, S, M и G1 циклины.
G1/S циклины (Cln1 и Cln2 у S. cerevisiae, циклин E у позвоночных) достигает
максимальной концентрации в поздней G1-фазе и падает в S-фазе.
G1/S cyclin–Cdk комплекс запускает начало репликации ДНК выключая различные
системы подавляющие S-phase Cdk в G1-фазе
G1/S циклины также инициируют дупликацию центросом у позвоночных, образование
веретенного тела у дрожжей. Падение уровня G1/S сопровождается увеличением
концентрации S циклинов (Clb5, Clb6 у Sc и циклин A у позвоночных), который
образует S циклин-Cdk комплекс который напрямую стимулирует ДНК репликацию.
Уровень S циклина остается высоким в течении всей S, G2-фаз и начала митоза,
где помогает началу митозу в некоторых клетках.
М-циклины (Clb1,2,3 и 4 у Sc, циклин B у позвоночных) появляется последним.
Его концентрация увеличивается, когда клетка переходит к митозу и достигает
максимума в метафазе. М-циклин-Cdk-комплекс включает сборку веретена деления
и выравнивание сестринских хроматид. Его разрушение в анафазе приводит
к выходу из митоза и цитокиезу.
G1 циклины (Cln3 у Sc и циклин D у позвоночных) помогает координировать
клеточный рост с входом в новый клеточный цикл. Они необычны, так как
их концентрация не меняется от фазы клеточного цикла, а меняется в ответ
на внешние регуляторные сигналы роста.
APC комплекс (Anaphase-Promoting Complex)
Убиквитин лигаза митоза - APC состоит из 12 субъединиц и регулирует
различные процессы митоза, такие как разделение сестринских хроматид (запускает
разрушение когезинов), переход к анафазе, анафазное расхождение хромосом,
выход из митоза, разрешение S-фазы. ARC разрушает митотический циклин
B.
Имеются различные белки регулирующие активность ARC комплекса, такие как
Mps1, Bub1, Bub3, BubR1, Mad1 и Mad2. Они ингибируют ARC комплекс, что
ведет к остановке клеточного цикла в метафазе митоза.
Ингибиторы комплексов циклин-Cdk
| Вид | Название | Синонимы | Гомологи | Цели, функции |
| S. cerevisiae | Sic1 Far1 |
Rum1 нет |
ингибирует S- и M-Cdk, подавляет активность Cdk в G1 ингибирует G1/S–Cdk в ответ на феромоны |
|
| S. pombe | Rum1 | Sic1 | ингибирует S– и М–Cdks, подавляет активность Cdk в G1 | |
| D. melanogaster | Roughex/Rux Dacapo/Dap |
нет Cip/Kip |
ингибирует S– и M–Cdk, подавляет активность Cdk in G1 ингибирует G1/S–Cdks, подавляет активность Cdk в G1 |
|
| X. laevis | Xic1 | Kix1 | Cip/Kip | ингибирует G1/S– и S–Cdk |
| H. sapiens | p21 p27 p57 p15INK4b p16INK4a p18INK4c p19INK4d |
Cip1/Waf1 Kip1 Kip2 - - - - |
Cip/Kip Cip/Kip i Cip/Kip INK4 INK4 INK4 INK4 |
ингибирует G1/S– и S–Cdk, активирует cyclin D–Cdk4 ингибирует G1/S– и S–Cdks, активирует cyclin D–Cdk4 ингибирует G1/S– и S–Cdks, активирует cyclin D–Cdk4 ингибирует Cdk4, Cdk6 ингибирует Cdk4, Cdk6 ингибирует Cdk4, Cdk6 ингибирует Cdk4, Cdk6 |
Помимо циклинов имеются и другие регуляторы, такие как p27, p53, анафазный
ингибитор, Cdc20, polo-like киназа, aurora киназа, Nek2 киназа и кинезинзависимые
моторные белки.
p53 (опухолевый супрессор) - нестабильный белок, узнает
поврежденную ДНК, стабилизируется, накапливается и стимулирует синтез
ингибитора Cdk2. Нарушения в работе p53 приводят к развитию раковых заболеваний.
p21 семейство - белки содержащие гомологичные N-концевые
участки, взаимодействующие с Cdk-циклинами. Активируются в стареющих клетках.
Образование индуцируется опухолевым супрессором p53. Может блокировать
субъединицы ДНК-полимеразы
p27 - межклеточные контакты стимулируют его синтез -
контактное подавление клеточного роста – задержка в G0. Когда
фибробласты помещаются в чашку они начинают делиться пока монослой не
покроет чашку – контактное торможение. В опытах показано что существенную
роль играют не контакты, а степень распластывания – чем меньше распластана
клетка, тем больше времени занимает клеточный цикл. Округление клеток
сопровождается снижением общей интенсивности белкового синтеза.
p57
p15, p16 семейство: взаимодействуют
с Cdk4 и Cdk6. Нарушают связь с циклинами D. Вероятно, задерживают рост
клеток.
Сокращения:
cdc-гены (cell-division-cycle genes) - гены клеточного цикла.
Литература
Suvar
June 21, 2006 19:17
Свои пожелания отправляйте по адресу
drosophila.narod.ru
Жизнь коротка, наука вечна
Процесс научного исследования глубоко интимен: иногда мы сами не знаем,
что мы делаем. (Ф.Крик)
![]()