Мембраны.
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
 рис.1
Плазматическая мембрана. Виден липидный бислой. Светлая область - внеклеточное
пространство, темная - внутриклеточное (Rees, 1996. P. 68).
рис.1
Плазматическая мембрана. Виден липидный бислой. Светлая область - внеклеточное
пространство, темная - внутриклеточное (Rees, 1996. P. 68).Моделью строения плазматической мембраны являестя жидкостно-мозаичная
модель (рис.1), обладающая свойствами замкнутости, текучести и асимметричности.
Замкнутость. Плазматическая мембрана является внешней
границей клетки, а также внутренних клеточных компартментов.
Текучесть. Липиды, белки и другие составляющие плазматической
мембраны движутся в пределах слоя. Переходы между слоями называются flip-flop,
происходят реже чем в пределах слоя, что обеспечивает наличие свойства
асимметричности. Переходы между слоями осуществляют ферменты транслокаторы
фосфолипидов
Асимметричность. Внешняя и внутренняя поверхности мембраны
различаются по составу липидов белков и наием гликокаликса на внешней
поверхности мембраны.
 рис.1
Жидкостно-мозаичная модель плазматической мембраны. Двойной слой липидов,
пронизанный различными белками. Наружная поверхность белков соединена
с сахарами, образующими гликокаликс. Липиды состоят из гидрофобной части
расположенной внутри бислоя (оранж.) и гидрофильной наружной части (сер.).
рис.1
Жидкостно-мозаичная модель плазматической мембраны. Двойной слой липидов,
пронизанный различными белками. Наружная поверхность белков соединена
с сахарами, образующими гликокаликс. Липиды состоят из гидрофобной части
расположенной внутри бислоя (оранж.) и гидрофильной наружной части (сер.).

мембраны газовых вакуолей некоторых бактерий 2 нм, состоят из белков
липидный бислой | мицелла, бислойная пленка-образования амфипатических
м-л в H2O | искусств бислои – сферические везикулы - липосомы, черные
м-ны – кажутся черными из-за интерференции света | липиды 50%m пм – фосфолипиды,
холестерол, гликолипиды – амфипатические м-лы – гидрофильная и гидрофобная
часть | Длина гидрофобного хвоста 14-24 С (всегда четно) | цис-ненасыщ
образ изгиб предающий рыхлость м-не | коэф. диффузии липидной м-лы D =
10-8 см2/с - ~2мкм/с | липиды вращ вокруг оси, хвосты гибкие | в миелине
~1500 разных липидов | белки:липиды=от1:4в миелине до 3:1 в бактериях,
липиды обеспеч электрич сопротивление, непрониц для полярных соед и прониц
для непол | м-ны способны образовывать отверстия ч-з кот проходят малые
мол-лы напр H2O | более короткие цепи упакованы в менее жесткие стр-ры
– уменьш вязкости м-ны, удлинение ацильной цепи уменьшает текучесть м-ны
| двойные связи повышает текучесть м-ны | повышение t повышает текучесть
м-ны | поля (patch)– мол-лы с разными св-вами могут объединяться в опред
месте напр жесткая часть пм прикрепляет кл к субстрату | вязкая м-на-более
проницаема для малых м-л (глюкоза) | замораживание скалывание:P-пов-ть
– гидрофоб часть внутр слоя м-ны, Е-пов-ть – гидрофоб часть внеш половины
бислоя
Компаненты мембран.
ГЛИКОЛИПИДЫ - липиды, содержащие олигосахарид.
Располагаются только в наружной мембране т.к. ферменты присоединяющие
сахар находятся внутри аппарата Гольджи. Гликокаликс -
мукополисахаридный наружный слой 3-10 нм в толщину, состаящий из остатков
олигосахаридов, имеет отрицательный заряд и составляет ~10% от массы мембраны.
Гликокаликс гидратирован, представляя собой желеподобную структуру.
Галактоцереброзид-основной нейтральный гликолипид миелиновой оболочки
аксонов нервных клеток.
Ганглиозиды - гликолипиды с остатком сиаловой кислоты (GM1 - связывает
холерный токсин).
 ХОЛЕСТЕРОЛ придает механическую прочность бислою, заполняя
свободное пространство, большую текучесть, способен свободно перетекать
из слоя в слой, обеспечивая изменение формы мембран при сжатии и растяжении.
ХОЛЕСТЕРОЛ придает механическую прочность бислою, заполняя
свободное пространство, большую текучесть, способен свободно перетекать
из слоя в слой, обеспечивая изменение формы мембран при сжатии и растяжении.
Мембраны прокариотических клеток не содержат холестерол.
ФОСФОЛИПИДЫ фосфоглицериды – глицерол,
две гидроксильные группы которого этерифицированы жирными кислотами, третья
фосфорной кислотой, этерифицированной спиртом*.
фосфатидилхолин - холиновая группа, *(CH3)3N+-CH2CH2OH
фосфатидилэтаноламин - *H2N-CH2CH2OH
фосфатидилсерин
кардиолипин – в значит кол-вах в м-нах бактерий и во внутр м-не
митох |
фосфатидилинозитол
cфингомиелин – отсутствует в мембране митохондрий.
фосфатидилглицерин - *глицерин. Является главным фосфолипидом
фотосинтетического аппарата всех растений. Содержится также в мембранах
сине-зеленых водорослей.
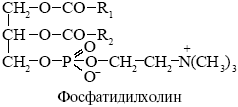
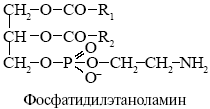
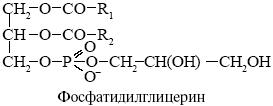
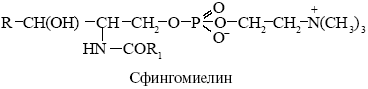
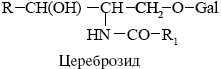
Фосфолипазы - ферменты разрушающие фосфолипиды в различных местах, используются для анализа мембранных липидов, некоторые получают из яда змей.
ИНТЕГРАЛЬНЫЕ БЕЛКИ.
Гликофорин А, содержащийся в мембранах эритроцитов, – первый изученный
интегральный белок. Содержит 24 аминокислоты в а-спирали представляют
собой трансмембранный домен:
arg+-val-gln-leu-ala-his-his-phe-ser-glu-pro-glu-ile-thr-leu-ile-ile-phe-gly-val-met-ala-gly-val-ile-gly-thr-ile-leu-leu-ile-ser-thr-gly-ile-arg+
Заряженные аминокислоты нейтрализуют друг друга His+ -Glu
Монотопные белки пересекают мембрану 1 раз, политопные белки - несколько
раз.
протеолипиды
протеогликаны
периферические белки – (спектрин- на внутр м-не, фибронектин-на внешней
м-не) удерживаются ионными взаимод, соединены с интегральными белками.
белки цитоскелета эритроцитов: полоса 1,2 спектрин (240-220кДа);
п2.1,2.2-анкирины(200-210кДа); п3-главный анионно-обменный белок (93кДа)-анионный
канал, обмен HCO3- на Cl- при выведении CO2; п4.1- белок связанный с о
спектрином и актином; п4.2-N-миристилиновый белок связан с белком п3 и
анкирином ; п4.9-актин-связывающий белок; п5- актин (43кДа).
Детергенты (SDS, тритонX-100) разрушают мембраны клеток. Внутренняя поверхность
мембраны эритроцитов заряжена отрицательно из-за содержания фосфатидилсерина.
Состав мембраны эритроцитов: гликолипиды-10%, холестерол-30%, фосфолипиды-60%:
фосфатидилхолин-28%, фосфатидилэтаноламин-27%, сфингомиелин-26%, фосфатидилсерин-13%.
контактное торможение роста (density-dependent inhibition, DDI) мутанты
образуют опухоли.
Литература.
Rees W.E. Life essay, 1996
Suvar
June 28, 2006 14:15
Свои пожелания отправляйте по адресу
drosophila.narod.ru
Жизнь коротка, наука вечна
Наука нам сокращает опыты быстротекущей жизни. ( А.С. Пушкин)
![]()